a LaFrance Consulting Services™ publication
TwoOldGuys™ Study Guides
Teacher Preparation for Biology Content Knowledge
by Charles R LaFrance, PhD
5. Ecology
Ecology is the study of the interactions among the organisms on a site, and of the
interactions between the organisms and the physical environment. The levels of
organization involved are the population, the community, and the ecosystem (and perhaps the
entire Biosphere, or all parts of the entire Earth where Life can be found). The ultimate
intent is to develop the necessary theories to account for the organizational dynamics of
the Biosphere. An important point here is that organization at ecological levels is
dynamic, or is subject to change over time. We are currently in a time period of Climate
Change, potentially a major Climate shift.
The controversy among Scientists is about the causes of this Climate Change, not
whether or not Climate Change is real [The magnitude of the Climate Change is a matter
of speculation, not of Science].
Given the challenges for Human Civilization to persist during the current Climate Change,
there is some urgency to understand how (or even if) Life as we know it can survive this
Climate Change. It is worth noting that Life itself can, and has, survived several major
Climate shifts (between global tropical conditions and global Ice Ages), but with mass
extinctions followed by complete replacement of the dominant Plants and Animals on Earth.
Understanding Nature becomes complicated because there are so many species involved,
even on rather small tracts of land: on enough undeveloped land (such as a weedlot) to hold
a typical suburban school (including the school grounds as well as the building) there
could be 100 or so plant species, 100’s of Arthropod species, and 10’s of
Chordate species.
If we hope to have our students begin to understand how Nature functions, we must start by
examining artificial, simplified models of natural systems of living creatures. Thus we
begin with the population.
5.1 Populations
In Ecological terms, a Population is
“a group of individuals of the same species occupying a single site
and interacting with each other.”
We have already seen two aspects of the
population in Chapter 3.2 Classical Genetics & Evolution, where we examined
Population Growth Potential and Population Genetics and the implications of these for the
concept of Evolution. In Genetics and Evolution, the most interesting interaction between
the members of the population is reproduction, as a means for transmitting genetic
information from one generation to the next. In Ecology, the more interesting interaction is
competition. We already know from Genetics and Evolution that competition must occur because
we can observe that populations change little from year to year in spite of the reproduction
potential to grow (in number of individuals) indefinitely.
There are five population parameters which we shall examine in this
chapter: Population density, Dispersion, Migratory patterns, Age
structure, and Distribution of populations.
Population density
The number of individuals in a population is called “population density.”
Density seems a reasonable term here because the population is defined as occupying a
given site, implying that it has a definable area. The details of the behavior of
population density were initially discovered in a series of laboratory experiments in
which the population could be isolated from other populations. The series of experiments
began with a single species, then added a second species. The analysis of the results led
to the development of tentative mathematical descriptions of population growth. However,
the math involved was Calculus which was designed by Isaac Newton specifically to explain
Physics (which he considered to behave as a rigorous mathematical system, with simple
cause-effect relationships). The assumptions of Calculus do not seem to apply well to
Ecology which appears to have multiple possible states (or effects), each with a
probability of occurring given a single cause.
Single species populations
When we compare the Reproductive potential (non-overlapping generations, litter
size = 6, from Chapter 3.2) to simulated data (environment [available food supply] will
support only 26 mice) for “actual,” observed mouse population in a house,
we see that the population does not grow as fast as its potential, due to competition. The
population even drops as the competition becomes too severe. It will recover over 3
generations back up to 9 pairs, then drop again. Thus one cause of fluctuations in the
population size is a somewhat cyclic rise and fall due to competition between members of
the population. Since this is typical of living systems (but less so for non-living,
chemical and physical, systems), the fluctuating population size is called a “dynamic
equilibrium.”
| House mice (Mus musculus) |
| |
potential |
population |
| |
date |
n |
pairs |
n |
pairs |
| P-1 |
9/22/10 |
2 |
1 |
2 |
1 |
| F-1 |
12/1/10 |
6 |
3 |
5 |
2 |
| F-2 |
2/9/11 |
18 |
9 |
9 |
4 |
| F-3 |
4/20/11 |
54 |
27 |
15 |
7 |
| F-4 |
6/29/11 |
162 |
81 |
17 |
8 |
| F-5 |
9/7/11 |
486 |
243 |
16 |
8 |
| F-6 |
11/16/11 |
1458 |
729 |
18 |
9 |
| F-7 |
1/25/12 |
4374 |
2187 |
15 |
7 |
Two species populations
Based on the definition of a population, it is not possible to have a population of two
species, so by ‘two species populations’ we clearly mean to say “two
populations.” A ‘species population’ uses ‘species’ (improperly)
as a adjective to distinguish the population from a statistical population, which is a
defined group of data points (the definition describes the statistical universe from which
the sample data can be drawn). The distinction between a species population and a
statistical population is needed because at least a majority of ecologists currently use
statistical methods to design experiments and to analyze the resulting data.
competition
When we add a second population, there is inter-species competition in addition to the
intra-species competition. The second species will make demands on the same resources over
which the members of the first species were already competing. At the dynamic equilibrium,
as one species declines, the other will grow, and the fluctuations in either species
population can be expected to be rather larger than in the artificial single population
model. By extension, we should not be surprised [although the environmentalists tend to
be not only surprised, but alarmed as if the natural world were tetering on the brink of
collapse] if the fluctuations increase even more as more species are added to the system.
These systems of multiple species are called Communities,
defined as “all populations (usually of species within the same
Order) occupying the same site.” Plant commuities of more than 200 species are
not unusual; Insect communities and Spider communities have more species than do Plant
communities; Amphibian and Reptile [called herpetological] communities, Bird communities,
and Mammal communities) communities generally have fewer species.
populations of predators and prey
A special case is when one population (predator) eats the other population (prey). In
such cases, there is no competition for the same resources; the prey species is the
resource for the predator. In such model systems under laboratory situations, an increase
in the prey species population results in more food for the predators, so the predator
population grows. The increase in predator population increases the number of prey eaten,
causing a decline in the prey population. The decline in prey population decreases the food
supply for the predators, causing a decline in the predator population. The decline in the
predator population decreases the numbers of prey eaten, causing the prey population to
increase. Thus the two populations exhibit strongly coupled cyclic fluctuations.
Adding more prey species allows the predators to ‘switch’
prey as the initial prey population declines; and adding more predator species causes
competition between predator species for the prey. In both the case of multiple prey species
and the case of multiple predator species, the coupling of the predator and prey species
fluctuations becomes weaker. It is believed that this leads to a damping of the flucuations
in the wild compared to what is seen in the laboratory. To my knowledge, there is no data
to support this hypothesis, and ample data to support the notion that fluctuations in
population density occur in natural systems.
Other Population Parameters
Dispersion describes the spacing between individuals of the population, and the
mechanisms utilized by the species to create and maintain this spacing. In Plant Ecology,
we consider “evenly dispersed” to mean that the individuals are randomly
distributed within the population; “clumped” refers to those species in which the
probability of finding another individual near an observed individual is higher than what
would occur if the individuals were evenly dispersed; and “over-dispersed” refers
to individuals maintaining an area of fewer individuals near an observed individual. Clumped
dispersions tend to occur when seeds travel only short distances from the parent plant, and
when asexual reproduction is commonplace. The most spectacular instances of over-dispersion
occur in species whose leaves release a growth-suppressing chemical as the leaves decompose
[for example creasote bush in Utah and California semi-deserts release creasote to suppress
seed germination reducing competition for limited water; creasote is highly flammable, and
burns with high temperature flames, leading to the spectacular wildfires of southern
California]. In Animal Ecology, there are behavioral patterns [such as territoriality, in
which a breeding pair will defend an area around the nest to preserve the food supply for
feeding the young]. “Home range” is the area in which the individuals search for
food, but without defending the home range from invaders (although many Animals
‘mark’ their home range with scents to discourage invaders).
Migratory patterns. “Migration” as used in Animal
Ecology implies the intent to return, and usually applies to seasonal movements between the
breeding season (Summer) grounds and the off-season (Winter) grounds, which may involve
North-South migrations or high altitude-low altitude migrations. When the young adults
migrate from their parents home range in search of breeding sites, they emigrate from one
area, and immigrate to another as a ‘permanent’ move. A mneumonic to remember
which is which: “Emigrate begins with ‘E’ as in EXIT while
Immigrate begins with ‘I’ as in IN.”
Age structure was introduced to predict future population
densities. The procedure required creating a “Life Table”
(lxmx-table) based on cohort (a group of individuals born during a
given time period, usually a year. The calculations involved are difficult (and confusing),
but not worth repeating here because the Life Table is rarely used today. I will show you
three examples just so you can get an idea of how this works.
The terms in the table are:
x = age group, usually a calendar year,
Nx = number alive at start of interval,
lx = proportion surving to end of interval,
dx = number dying during interval,
qx = probability of dying, or age-specific mortality,
ex = life expectancy for survivors, as the number of intervals that
50% of the individuals in the current age group can be expected to survive.
Example 1. A full Life Table for a population of Red
deer, a wild animal found in southern England.
Red Deer females
Lowe, 1969 |
| x |
Nx |
lx |
dx |
qx |
ex |
| 2 |
1000 |
1.000 |
61 |
0.0610 |
3.35 |
| 3 |
939 |
0.939 |
185 |
0.1970 |
2.53 |
| 4 |
754 |
0.754 |
249 |
0.3302 |
2.03 |
| 5 |
505 |
0.505 |
200 |
0.3960 |
1.79 |
| 6 |
305 |
0.305 |
119 |
0.3901 |
1.63 |
| 7 |
186 |
0.186 |
54 |
0.2903 |
1.35 |
| 8 |
132 |
0.132 |
107 |
0.8105 |
0.70 |
| 9 |
25 |
0.025 |
25 |
1.0000 |
0.50 |
Example 2. an abbreviated Life Table for a population of
Rhodendron sp, a wild plant found in the Smokey Mountains of the U.S. southeast
Rhododendron sp
McGraw, 1989 |
| x |
|
lx |
dx |
qx |
ex |
| 0 |
|
1.000 |
0 |
0 |
5.60 |
| 1 |
|
1.000 |
0.016 |
0.016 |
4.60 |
| 2 |
|
0.984 |
0 |
0 |
3.67 |
| 3 |
|
0.984 |
0.075 |
0.077 |
2.67 |
| 4 |
|
0.909 |
0.185 |
0.024 |
1.85 |
| 5 |
|
0.724 |
0.346 |
0.477 |
1.19 |
| 6 |
|
0.378 |
0.270 |
0.714 |
0.82 |
| 7 |
|
0.108 |
0.095 |
0.882 |
0.62 |
| 8 |
|
0.013 |
0.013 |
1.000 |
0.50 |
Example 3. a highly abbreviated Life Table for the 1980
Human population of the United States (U.S. Census Bureau data). The interval is one year,
but only every 10th year is shown to shorten the table. The current
full table can be viewed at
2011 Statistical Abstracts, U.S. Census Bureau.
Life expectancy
Human, USA 1980 |
|
male |
female |
| x |
ex |
ex |
| 0 |
70.83 |
75.83 |
| 10 |
61.66 |
66.53 |
| 20 |
52.37 |
57.04 |
| 30 |
43.24 |
47.65 |
| 40 |
34.05 |
38.36 |
| 50 |
25.36 |
29.53 |
| 60 |
17.51 |
21.25 |
| 70 |
10.96 |
13.67 |
| 80 |
6.18 |
7.48 |
| 90 |
3.18 |
3.45 |
| 100 |
0.50 |
0.50 |
If you were a 30 year old male in 1980, you could have expected to live another 43
years and about 3 months or to the age of 73. If you were a 30 year old female in 1980,
you could have expected to live another 47 years and about 6 months or to the age of 77.
More accurately, of all 30 year old males alive in 1980, 50% would have been expected to
die before the age of 73 yrs 3 months, and the other 50% would have been expected to live
longer than an additional 43 yrs 3 months; and of all 30 year old females alive in 1980,
50% would have been expected to die before the age of 77 yrs 6 months, and the other 50%
would have been expected to live longer than an additional 47 yrs 6 months. For what it
is worth, out of 10,000 30yr old men buying $20,000 simple life insurance policies in
1980, the insurance company would expect to pay out $20,000 to 5,000 policy holders or
$100,000,000 by 2023, so would need to collect $232.56 per year (or $19.38 per month) from
each of the 10,000 policy holders to break even [this is the legal definition of simple
life insurance].
The insurance company would profit on the investment of the net collected premiums until
the last surviving policy holder died. Each surviving policy holder would pay the $232.56
per year until death. Of course, the insurance company gets to keep all collected premiums
on each policy which lapsed due to non-payment of the annual premiums.
Distribution of populations
Most populations, and species, have limited distribution. An extreme of limited
distribution is Kirkland's warbler (Dendroica kirklandii, Songbird Family) with a
breeding range including only sandy jack pine barrens of northern lower Michigan, or about
15,500 km2 (6,000 sq miles) or 1,600,000 hectares (3,800,000 acres). This
illustrates the two aspects of distribution: geographic limits [northern lower Michigan]
(often stated as “county records” or a list of all counties where specimens have
been collected or observed by reliable observers) and habitat
(“a general description of the environmental conditions where the
population can be expected to be present”) [sandy Jack pine barrens]. For any
of the readers who are interested, Kirkland's warbler winters in the Bahamas and nearby
islands (Turks, Caicos and Hispaniola).
In 1840, Leibig published the Law of the Minimum [Sprengel actually
publish the same hypothesis in 1828] which hypothesizes that, for Plants, one of the
factors in the physical-chemical environment will limit the growth of a plant (and if the
amount of the limiting factor is increased, another factor will become limiting). For
Animals, the limiting factor will usually be the food supply. The Sprengel-Leibig Law is
still used in agriculture to design fertilizer regimes to increase yields.
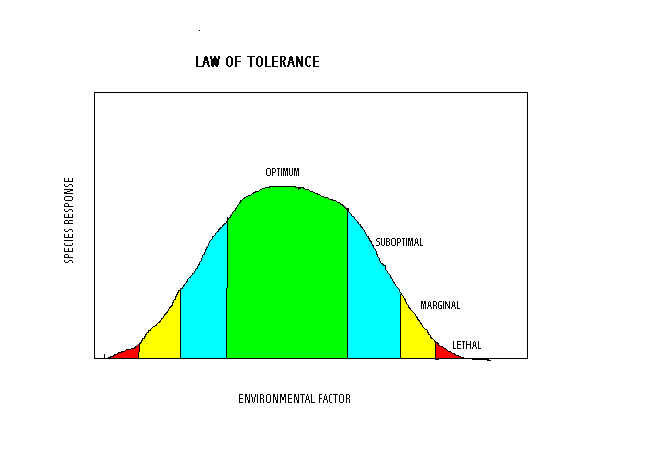
The principle of limiting factors was generalized by Shelford in 1911 as the Law of
(Ecological) Tolerance. As now understood (after research on the physiological response of
plants to the levels of environmental factors), Ecological tolerance is a bell-shaped curve
(probably the statistical Normal distribution familiar to teachers as the grading curve)
showing the relationship of plant response to environmental factors. The highest point on
the curve is the optimum condition at which the plant has the best growth, reproduction,
etc. Shelford divided the rest of the curve into ‘zones:’ an
optimal zone [the “C” grades] on either side of the optimum point where growth
is good; sub-optimal zones on either side of the optimal zone (the “B” and
“D” grades]; beyond the sub-optimal zones are the marginal zones [“A”
and “F” grades] where survival is barely possible; and the lethal zones where the
plant cannot survive. If we knew the physiological response of the species to the
environmental factors involved, we should be able to predict how well the population would
do under any level of the environmental factor. When the environment forms a gradient, such
as depth to the water table (and therefore availability of water, or risk of flooding) from
a stream, across the floodplain and up into the surrounding hills, this model predicts that
the population density will change along the gradient. Other species populations on the site
should, according to modern interpretation of the theory of Evolution, have different optimum
points. This is ‘plot material’ for the next two sections (Chapter 5.2
Communities, and Chapter 5.3 Ecosystems).
Works Cited
RETURN to
TwoOldGuys HOME
OUTLINE, Biology: Content Knowledge
© 2004-2011 TwoOldGuys ™
revised: 19 Jul 2011